Zgodnie z teorią Morgana geny zajmują ściśle zdefiniowane miejsce na chromosomie tzw. loci. Geny znajdujące się na jednym chromosomie określane są genami sprzężonymi lub inaczej współdziedziczonymi, ponieważ podczas podziałów komórkowych do komórek potomnych przemieszczają się całe chromosomy, a nie ich fragmenty. Zaburzeniem tego zjawiska jest mechanizm crossing-over, który polega na wymianie odcinków chromatyd pomiędzy chromosomami homologicznymi w czasie mejozy, co zwiększa różnorodność genetyczną i "osłabia" sprzężenie ze sobą genów. Co więcej, to dzięki crossing-over możliwe jest określenie odległości pomiędzy dwoma wybranymi genami obecnymi na jednym chromosomie i tworzenie ich map.
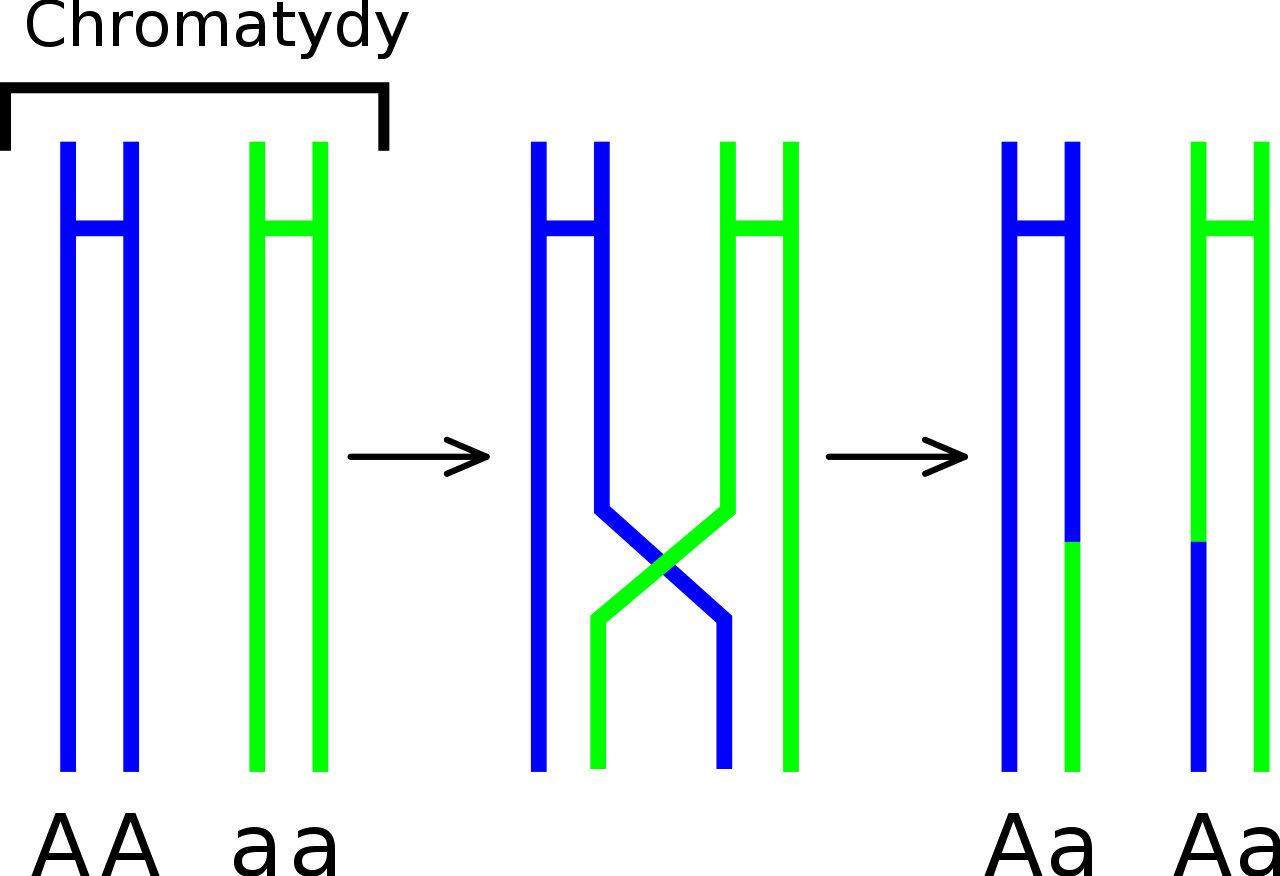
Schemat crossing-over - zachodzi wymiana alleli pomiędzy chromatydami
Prawdopodobieństwo zajścia crossing-over pomiędzy dwoma wybranymi genami jest tym większe, im większa jest odległość między nimi — dłuższy odcinek pomiędzy genami gwarantuje więcej miejsc, gdzie crossing-over może zajść. Tym samym geny leżące blisko siebie na chromosomie można określić jako silniej sprzężone, ponieważ istnieje mała szansa, że to właśnie pomiędzy nimi zajdzie wymiana odcinków w procesie crossing-over. Na podstawie badań nad krzyżowaniem czystych linii muszek owocowych, Morgan opracował jednostkę mapową (j.m. lub centymorgan cM), która definiuje odległości pomiędzy genami. Jest ona oparta na licznie rekombinantów wśród potomstwa - 1% zrekombinowanych osobników w pierwszym pokoleniu krzyżówki osobników czystej linii odpowiada 1 jednostce mapowej. Do tworzenia istotnych statystycznie map chromosomowych potrzebna jest duża liczba potomstwa, dlatego muszka owocowa była idealnym modelem dla potrzeb Morgana. Wzór na jednostkę mapową można zapisać w postaci:

Określenie rekombinatów, dotyczy osobników, które posiadają inny (zrekombinowany) zestaw cech niż pokolenie rodzicielskie — świadczy to o zajściu crossing-over pomiędzy genami kodującemi te cech (o ile występują na jednym chromosomie).